��Ŀ�� ��Դ�� �}�ͣ�����}
�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ�����}
�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ�����}
�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ�����}
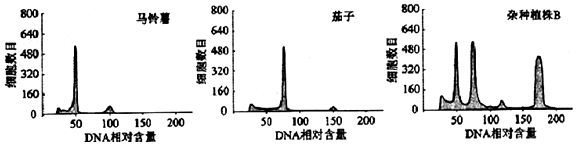
�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ�����}

�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ�����}
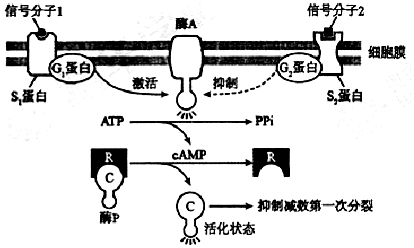
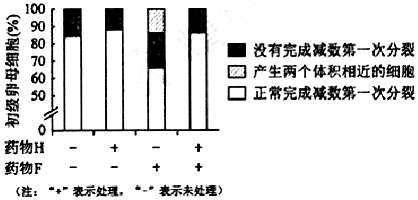
�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ�����}
 �����ˆT�о����չ�����е��S���~Ƭ�Ĺ�����ã�
�����ˆT�о����չ�����е��S���~Ƭ�Ĺ�����ã�| �~λ | ������ȣ���m�� | Ƭ�Ӕ�(sh��) |
| ��λ�~ | 1.79 | 10.90 |
| ����λ�~ | 2.46 | 17.77 |
| �����~ | 3.06 | 17.91 |
�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ�����}

�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ��x���}
| A�� | ���B(y��ng)ԭ�����Ǽ�����ȫ���� | B�� | ��ͨ�^�нz�������Ӽ�����(sh��)Ŀ | ||
| C�� | ���B(y��ng)���϶����ȡ�Է����M�� | D�� | ����(j��ng)�^Ó�ֻ��֏�(f��)����ȫ���� |
�鿴�𰸺ͽ���>>
��Ŀ�� ��Դ�� �}�ͣ��x���}
| A�� | �����ȵ���ø̎����Ҭ�˺ͺڽ�@��ԭ���|(zh��)�w | |
| B�� | Ⱦɫ�w��(sh��)��26����Ʒ�Nһ���������w | |
| C�� | ������Ʒ�N�Ŀ���������܁��Բ�ͬȾɫ�w | |
| D�� | ������Ʒ�N��Ҭ�˾������γɿ������� |
�鿴�𰸺ͽ���>>
�ٶ����� - ����(x��)���б� - ԇ�}�б�
����ʡ��(li��n)�W(w��ng)�`���Ͳ�����Ϣ�e��ƽ�_ | �W(w��ng)���к���Ϣ�e�^(q��) | ����p�_�e�^(q��) | ��vʷ̓�o���x�к���Ϣ�e�^(q��) | �����֙�(qu��n)�e�^(q��)
�`���Ͳ�����Ϣ�e���Ԓ��027-86699610 �e���]�䣺58377363@163.com